本试题 “对下列四幅图的描述,正确的是[ ]A.图甲可依次表示果蝇的一个精原细胞在减数分裂过程中等位基因分离时的染色单体数、DNA数、同源染色体对数B.图乙表示两种...” 主要考查您对自由组合定律
减数分裂
神经冲动的产生和传导
等考点的理解。关于这些考点您可以点击下面的选项卡查看详细档案。
- 自由组合定律
- 减数分裂
- 神经冲动的产生和传导
基因的自由组合定律与应用:
1.自由组合定律:控制不同性状的遗传因子的分离和组合是互不干扰的;在形成配子时,决定同一性状的成对的遗传因子彼此分离,决定不同性状的遗传因子自由组合。
2. 实质
(1)位于非同源染色体上的非等位基因的分离或组合是互不干扰的。
(2)在减数分裂过程中,同源染色体上的等位基因彼此分离的同时,非同源染色体上的非等位基因自由组合。
3.适用条件
(1)有性生殖的真核生物。
(2)细胞核内染色体上的基因。
(3)两对或两对以上位于非同源染色体上的非等位基因。
4.细胞学基础:基因的自由组合定律发生在减数第一次分裂后期。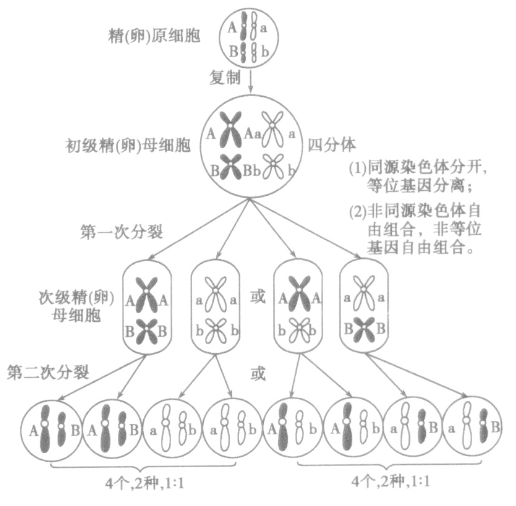
5.应用
(l)指导杂交育种,把优良性状重组在一起。
(2)为遗传病的预测和诊断提供理沦依据。
两对相对性状的杂交实验:
1.提出问题——纯合亲本的杂交实验和F1的自交实验
(1)发现者:孟德尔。
(2)图解:

2.作出假设——对自由组合现象的解释
(1)两对相对性状(黄与绿,圆与皱)由两对遗传因子(Y与y,R与r)控制。
(2)两对相对性状都符合分离定律的比,即3:1,黄:绿=3:1,圆:皱=3:1。
(3)F1产生配子时成对的遗传因子分离,不同对的遗传因子自由组合。
(4)F1产生雌雄配子各4种,YR:Yr:yR:yr=1:1:1:1。
(5)受精时雌雄配子随机结合。
(6)F2的表现型有4种,其中两种亲本类型(黄圆和绿皱),两种新组合类型(黄皱与绿圆)。黄圆:黄皱:绿圆:绿皱=9:3:3:1
(7)F2的基因型有16种组合方式,有9种基因型。
3.对自由组合现象解释的验证
(1)方法:测交。
(2)预测过程:

(3)实验结果:正、反交结果与理论预测相符,说明对自由组合现象的解释是正确的。
自由组合类遗传中的特例分析9:3:3:1的变形:
9:3:3:1是独立遗传的两对相对性状自由组合时出现的表现型比例,题干中如果出现附加条件,则可能出现9:3:4、9:6:1、15:1、9:7等一系列的特殊分离比。
特殊条件下的比例关系总结如下:
| 条件 | 种类和分离比 | 相当于孟德尔的分离比 |
| 显性基因的作用可累加 | 5种,1:4:6:4:1 | 按基因型中显性基因个数累加 |
| 正常的完全显性 | 4种,9:3:3:1 | 正常比例 |
| 只要A(或B)存在就表现为同一种,其余正常为同一种,其余正常表现 | 3种,12:3:1 | (9:3):3:1 |
| 单独存在A或B时表现同一种,其余正常表现 | 3种,9:6:1 | 9:(3:3):1 |
| aa(或hb)存在时表现为同一种,其余正常表现 | 3种,9:3:4 | 9:3:(3:1) |
| A_bb(或aaB_)的个体表现为一种,其余都是另一种 | 2种,13:3 | (9:3:1):3 |
| A、B同时存在时表现为同一种,其余为另一种 | 2种,9:7 | 9:(3:3:1) |
| 只要存在显性基因就表现为同一种 | 2种,15:1 | (9:3:3):1 |
注:利用“合并同类项”巧解特殊分离比
(1)看后代可能的配子组合,若组合方式是16种,不管以什么样的比例呈现,都符合基因自由组合定律。
(2)写出正常的分离比9:3:3:1。
(3)对照题中所给信息进行归类,若后代分离比为 9:7,则为9:(3:3:1),即7是后三种合并的结果;若后代分离比为9:6:1,则为9:(3:3):1;若后代分离比为15:1 则为(9:3:3):1等。
表解基因的分离定律和自由组合定律的不同:
| 分离定律 | 自由组合定律 | ||
| 两对相对性状 | n对相对性状 | ||
| 相对性状的对数 | 1对 | 2对 | n对 |
| 等位基因及位置 | 1对等位基因位于1对同源染色体上 | 2对等位基因位于2对同源染色体上 | n对等位基因位于n对同源染色体上 |
| F1的配子 | 2种,比例相等 | 4种,比例相等 | 2n种,比例相等 |
| F2的表现型及比例 | 2种,3:1 | 4种,9:3:3:1 | 2n种,(3:1)n |
| F2的基因型及比例 | 3种,1:2:1 | 9种,(1:2:1)2 | 3n种,(1:2:1)n |
| 测交后代表现型及比例 | 2种,比例相等 | 4种,比例相等 | 2n种,比例相等 |
| 遗传实质 | 减数分裂时,等位基因随同源染色体的分离而分开,分别进入不同配子中 | 减数分裂时,在等位基因随同源染色体分开而分离的同时,非同源染色体上的非等位基因自由组合,进而进入同一配子中 | |
| 实践应用 | 纯种鉴定及杂种自交纯合 | 将优良性状重组在一起 | |
| 联系 | 在遗传中,分离定律和自由组合定律同时起作用:在减数分裂形成配子时,既有同源染色体上等位基因的分离,又有非同源染色体上非等位基因的自由组合 | ||
易错点拨:
1、F2共有16种组合方式,9种基因型,4种表现型,其中双显(黄圆):一显一隐(黄皱):一隐一显(绿圆):双隐(绿皱)=9:3:3:1。F2中纯合子4种,即YYRR、YYrr、yyRR、yyrr,各占总数的 1/16;只有一对基因杂合的杂合子4种,即YyRR、Yyrr、 YYRr、VyRr,各占总数的2/16;两对基因都杂合的杂合子1种,即YyRr,占总数的4/16。
2、F2中双亲类型(Y_R_十yyrr)占10/16。重组类型占6/16(3/16Y_rr+3/16yyR_)。
3、 减数分裂时发生自由组合的是非同源染色体上的非等位基因,而不是所有的非等位基因。同源染色体上的非等位基因,则不遵循自由组合定律。
4、用分离定律解决自由组合问题
(1)基因原理分离定律是自由组合定律的基础。
(2)解题思路首先将自由组合定律问题转化为若干个分离定律问题。在独立遗传的情况下,有几对基因就可以分解为几个分离定律问题。如AaBb×Aabb可分解为:Aa× Aa,Bb×bb。然后,按分离定律进行逐一分析。最后,将获得的结果进行综合,得到正确答案。
知识拓展:
1、两对相对性状杂交试验中的有关结论
(1)两对相对性状由两对等位基因控制,且两对等位基因分别位于两对同源染色体。
(2)F1减数分裂产生配子时,等位基因一定分离,非等位基因(位于非同源染色体上的非等位基因)自由组合,且同时发生。
(3)F2中有16种组合方式,9种基因型,4种表现型,比例9:3:3:1。
2、自由组合定律的实质:减I分裂后期等位基因分离,非等位基因自由组合。
3、孟德尔成功的原因分析
(1)正确选择实验材料豌豆适合作杂交实验材料酌优点有:
①具有稳定的、易于区分的相对性状。
②严格自花传粉,闭花受粉,在自然状态下均是纯种。
③花比较大,易于做人工杂交实验。
(2)精心设计实验程序
①采取单一变量分析法,即分别观察和分析在一个时期内的一对相对性状的差异,最大限度地排除各种复杂因素的干扰。
②遵循了由简单到复杂的原则,即先研究一对相对性状的遗传,再研究多对相对性状的遗传,由此从数学统计中发现遗传规律。
③运用了严密的假说—演绎法。针对发现的问题提出假说,并设计实验验证假说,在不同的杂交实验中分别验证假说的正确性,从而使假说变成普遍的规律。
(3)精确的统计分析通过对一对相对性状、两对相对性状杂交实验中子代出现的性状进行分类、计数和数学归纳,找出实验显示出来的规律性,并深刻的认识到比例中所隐藏的意义和规律。
(4)首创了测交的方法巧妙地设计了测交方法,证明了假说的正确性。这种以杂交子一代个体与隐性纯合子进行测交的方法,已成为遗传学分析的经典方法。
4、遗传规律的再发现
(1)1909年,丹麦生物学家约翰逊把“遗传因子”叫做基因。
(2)因为孟德尔的杰出贡献,他被公认为“遗传学之父”。
例 下列哪项不是孟德尔选用豌豆作实验材料并获得成功的原因( )
A.豌豆具有稳定的、容易区分的相对性状
B.豌豆是严格闭花受粉的植物
C.用统计学的方法引入对实验结果的分析
D.豌豆在杂时,母本不需去雄
思路点拨:孟德尔获得成功的原因有:①正确选择实验材料;②精心设计实验程序;③精确的统计分析; ④首创了测交方法。孟德尔的杂交实验中,母本是必须去雄的。所以D选项错误。答案D
1、减数分裂概念的理解:
(1)范围:进行有性生殖的生物。
(2)在原始生殖细胞(精原细胞或卵原细胞)发展成为成熟生殖细胞(精子或卵细胞)过程中进行的。
(3)过程:减数分裂过程中染色体复制一次细胞连续分裂两次。
(4)结果:新细胞染色体数减半。
2、减数分裂中染色体的变化过程
复制→联会→四分体→同源染色体彼此分离→染色单体彼此分离。
(1)精子的形成过程:

(2)卵细胞的形成过程:
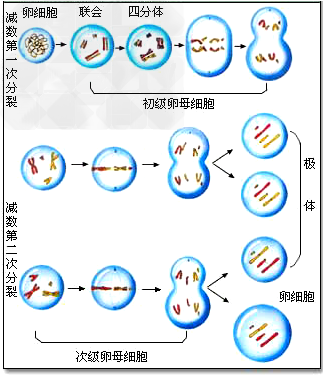
与减数分裂相关的概念辨析:
1.染色体和染色单体
(1)染色体的条数等于着丝点数,有几个着丝点就有几条染色体。
(2)染色单体是染色体复制后,一个着丝点上连接的两条相同的单体,这两条单体被称为姐妹染色单体,其整体是一条染色体。
2.同源染色体与非同源染色体
(l)同源染色体
①在减数分裂过程中进行配对的两条染色体。
②形状大小一般相同,一条来自父方一条来自母方。
(2)非同源染色体:在减数分裂过程中不进行配对的染色体,它们形状大小一般不同。
3.姐妹染色单体和非姐妹染色单体
(1)姐妹染色单体:同一着丝点连着的两条染色单体。
(2)非姐妹染色单体:不同着丝点连接着的两条染色单体。同源染色体内的非姐妹染色单体四分体时期可发生交叉互换。
4.联会和四分体
(1)联会:是指减数第一次分裂过程中(前期)同源染色体两两配对的现象。
(2)四分体:联会后的每对同源染色体含有四条染色单体,叫做四分体,即1个四分体=1对同源染色体 =2条染色体=4条染色单体=4个DNA分子。
细胞不同分裂图像的判断方法:
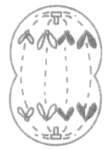
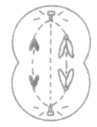
1.减数分裂和有丝分裂的不同分裂时期图像
| 前期 | 中期 | 后期 | ||
| 有丝分裂 | 有同源染色体(染色体必定成对) |  不联会 |
 着丝点排列在赤道板 |
 着丝点分裂 |
| 减数第一次分裂 | 有同源染色体(染色体必定成对) |  联会 |
 四分体排列在赤道板两侧 |
 同源染色体分离,非同源染色体自由组合 |
| 减数第二次分裂 | 无同源染色体(染色体可以不成对) |  无同源染色体 |
 着丝点排列在赤道板 |
 着丝点分裂 |
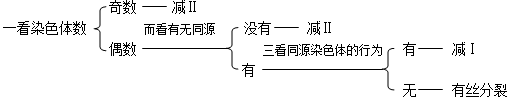
3.一般来说,细胞分裂方式的识别还需要观察细胞外形和细胞质的分裂方式等。
(1)细胞板隔裂——植物细胞分裂;细胞膜缢裂动物细胞分裂。
(2)矩形有壁为植物细胞;圆形无壁一般为动物细胞。
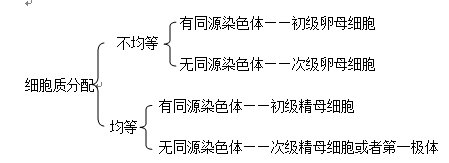
(3)依据细胞质分裂方式的判断:
表解减数分裂与有丝分裂的异同:
| 项目 | 有丝分裂 | 减数分裂 | |
| 不同点 | 母细胞 | 体细胞或原始生殖细胞 | 原始生殖细胞 |
| 染色体复制 | 一次,有丝分裂间期 | 一次,减数第一次分裂前的间期 | |
| 细胞分裂次数 | 一次 | 两次 | |
| 同源染色体行为 | 有同源染色体,但无同源染色体的联会、四分体,不出现同源染色体的非姐妹染色单体间的交叉互换及同源染色体的分离 | 有同源染色体的联会、四分体,出现同源染色体的非姐妹染色单体间的交叉互换及同源染色体的分离 | |
| 子细胞的数目 | 2个 | 雄性4个,磁性为(1+3)个 | |
| 子细胞的类型 | 体细胞 | 生殖细胞(精细胞或卵细胞) | |
| 最终产生子细胞染色体数 | 与亲代细胞相同 | 比亲代细胞减少一半(发生在减数第一次分裂) | |
| 子细胞核的遗传物质组成 | 几乎相同 | 不一定相同(基因重组形成多种配子) | |
| 子细胞中可遗传变异的来源 | 基因突变、染色体变异 | 基因突变、基因重组、染色体变异 | |
| 染色体及DNA数量的变化 |  |
 | |
| 相同点 | 染色体都复制一次;出现纺锤体,均有子细胞产生,均有核膜、核仁的消失与重建过程;减数第二次分裂和有丝分裂相似,着丝点分裂,姐妹染色单体分开形成染色体 | ||
| 意义 | 使生物的亲代和子代之间保持了遗传特性的稳定性 | 减数分裂和受精作用维持了每种生物前后代体细胞中染色体数目的恒定 | |
易错点拨:
1、同源染色体并非完全相同:同源染色体形态,大小一般都相同,但也有大小不相同的,如男性体细胞中X染色体和Y染色体是同源染色体, X染色体较大,Y染色体较小。
2、并非所有细胞分裂都存在四分体:联会形成四分体是减数第一次分裂特有的现象,有丝分裂过程中不进行联会,不存在四分体。
3、存在染色单体的时期是染色体复制之后、着丝点分裂之前的一段时期,如有丝分裂前、中期,减数第一次分裂,减数第二次分裂前、中期。
4、减数分裂过程中是先复制后联会,联会的两条染色体一定是同源染色体。联会是细胞进行正常减数分裂的基础。
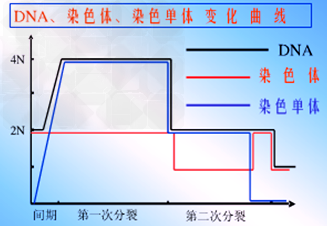
5、减数分裂过程中,染色体复制一次,细胞分裂两次,着丝点分裂一次,染色体减半一次,DNA减半两次。
6、原始生殖细胞染色体数目与体细胞相同,其既能进行有丝分裂产生原始生殖细胞,也能进行减数分裂。 7、以二倍体生物为例,具有同源染色体的细胞:精(卵)原细胞、初级精(卵)母细胞、体细胞。无同源染色体的细胞:次级精(卵)母细胞、精(卵)细胞、第一(二)极体、精子。
例 某动物的精子中有染色体16条,则在该动物的初级精母细胞中存在染色体数、四分体数、染色单体数、DNA分子数、脱氧核苷酸链数分别是( )
A,32、16、64、64、128
B.32、8、32、64、128
C.16、8、32、32、64
D.16、0、32、32、64
思路点拨:精子是精原细胞经过两次分裂得到的,其染色体数是精原细胞的一半,因此精原细胞中有同源染色体16对,初级精母细胞是精原细胞复制产生的,1条染色体含2个染色单体,2个DNA,4条DNA单链。答案A
减数分裂过程中DNA、染色体、染色单体的数目变化曲线:
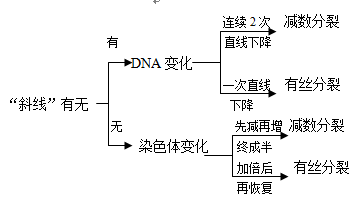
DNA与染色体的变化曲线识别方法
知识拓展:
1、AaBb(两对基因独立遗传)产生配子情况
| 产生的配子种类 | 说明 | |
| 一个精原细胞 | 4个,2种(AB、ab或Ab、aB) | 植物产生配子的结论,与动物大致相同(区别仅在于:一个花粉母细胞产生的精子数量是8个,比动物要多) |
| 一个雄性个体 | 4n个,4种(AB、ab、Ab、aB) | |
| 一个卵原细胞 | 1个,1种(AB或ab或Ab或aB) | |
| 一个雌性个体 | n个,4种(AB、ab、Ab、aB) |
①如果在四分体时期,不发生非姐妹染色单体的交叉互换,则一个精原细胞形成4个、2种精子细胞。
②如果在四分体时期,发生了非姐妹染色单体的交叉互换,则一个精原细脆形成4个、4种精子细胞。
③若两个精细胞中染色体完全相同,则它们可能来自同一个次级精母细胞。
④若两个精细胞中染色体恰好“互补”,则它们可能来自同一个初级精母细胞分裂产生的两个次级精母细胞。
⑤若两个精细胞中的染色体有的相同,有的互补,只能判定可能来自同一个生物不同精原细胞的减数分裂过程。
例 基因型为AaBb(两对等位基因位于两对同源染色体上)的一个精原细胞经减数分裂形成的精细胞种类有 ( ) A.1种 B.2种 C.4种 D.8种
思路点拨:一个基因型为AaBb的精原细胞减数分裂只形成2种类型的4个精子。但若一个基因型为AaBb个体产生的精子种类就应是4种。答案B
3、减数第一次分裂和有丝分裂的区别是前者同源染色体变化为四分体,后者不形成四分体。
4、减数第二次分裂同有丝分裂的区别是减数第二次分裂的细胞无同源染色体,而有丝分裂有同源染色体,其染色体行为的变化二者是相同的。
5、细胞分裂与变异类型的关系
| 分裂方式 | 变异类型 |
| 无丝分裂 | 基因突变 |
| 有丝分裂 | 基因突变、染色体变异 |
| 减数分裂 | 基因突变、基因重组、染色体变异 |
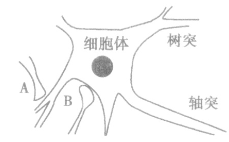
1、神经元:是一种高度特化的细胞,是神经系统的基本结构和功能单位之一,它具有感受刺激和传导兴奋的功能。神经细胞呈三角形或多角形,可以分为树突、轴突和胞体这三个区域。

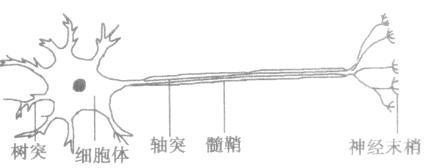
2、神经元按照用途分为三种:传入神经,传出神经,和连体神经。
3、根据神经元的功能又可分:
①感觉神经元(sensoryneuron),或称传入神经元(afferentneuron)多为假单极神经元,胞体主要位于脑脊神经节内,其周围突的末梢分布在皮肤和肌肉等处,接受刺激,将刺激传向中枢。
②运动神经元(motorneuron),或称传出神经元(efferentneuron)多为多极神经元,胞体主要位于脑、脊髓和植物神经节内,它把神经冲动传给肌肉或腺体,产生效应。
③中间神经元(interneuron),介于前两种神经元之间,多为多极神经元。动物越进化,中间神经元越多,人神经系统中的中间神经元约占神经元总数的99%,构成中枢神经系统内的复杂网络。
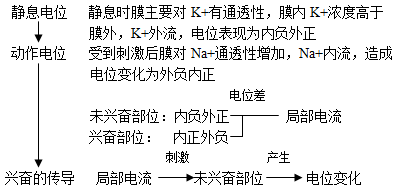
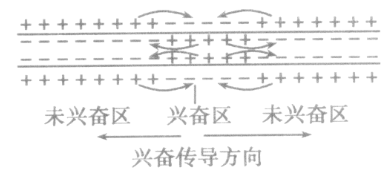
4、兴奋在神经纤维上的传导:以电信号的形式沿着神经纤维的传导是双向的;
静息时膜内为负,膜外为正(外正内负);
兴奋时膜内为正,膜外为负(外负内正),兴奋的传导以膜内传导为标准。
5、兴奋在神经元之间的传递——突触
(1)突触的结构:
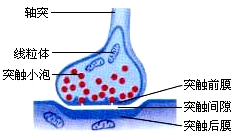
突触小体中有突触小泡,突触小泡中有神经递质,神经递质只能由突触前膜释放到突触后膜,使后膜产生兴奋(或抑制)所以是单向传递。(突触前膜→突触后膜,轴突→树突或胞体)
(2)从结构上来说,突触可以分为两大类:
A.轴突——树突表示为:

B.轴突——细胞体表示为:

(3)兴奋在神经元之间的传递
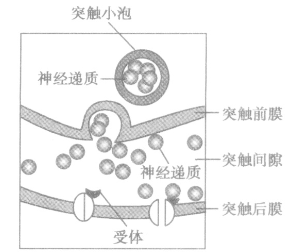
a、神经递质移动方向:突触小泡→突触前膜(释放递质)→突触间隙→突触后膜。
b、神经递质的种类:乙酰胆碱、多巴胺等。
c、神经递质的去向:迅速地分解或被重吸收到突触小体或扩散离开突触间隙,为下一次兴奋做好准备。
d、受体的化学本质为糖蛋白。
e、神经递质的释放过程体现了生物膜的结构特点——流动性。
f、传递特点:单向性,神经递质只能由突触前膜释放作用于突触后膜,引起下一个神经元的兴奋或抑制。
g、在突触传导过程中有电信号→化学信号→电信号的过程,所以比神经元之间的传递速度慢。
兴奋在神经纤维上的传导与突触传递的比较:
| 过程 | 特点 | 速度 | |
| 神经纤维 | 刺激→电位差→局部电流→未兴奋区 | 双向传导 | 快 |
| 神经细胞间 | 突触小体→突触小泡→递质→突触(前膜→间隙→后膜)→下一个神经细胞的树突或细胞体 | 单向传递 | 慢 |
知识点拨:
1、在膜外,兴奋传导方向与局部电流方向相反。在膜内,兴奋传导方向与局部电流方向相同。
2、在一个神经元内有一处受到刺激产生兴奋,迅速传至整个神经元细胞,即在该神经元的任何部位均可测到生物电变化。
3、判断突触前膜、突触后膜的方法已知突触结构图时,膜内有突触小泡,则该膜为突触前膜,否则为后膜。
4、神经递质作用效果有两种:促进或抑制。递质释放的过程为胞吐,由突触后膜(下一神经元的细胞体或树突部分)糖蛋白识别。
知识拓展:
一、电流表指针偏转方向与次数的判断
1.在神经纤维上
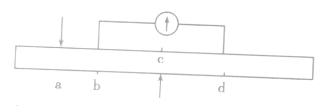
(1)刺激a点,b点先兴奋,d点后兴奋,电流计发生两次方向相反的偏转。
(2)刺激c点(bc=cd),b点和d点同时兴奋,电流计不发生偏转
2.在神经元之间
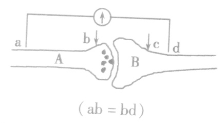
(1)刺激b点,由于兴奋在突触间的传递速度小于在神经纤维上的传导速度,a点先兴奋,d点后兴奋,电流计发生两次方向相反的偏转。
(2)刺激c点,兴奋不能传至a点,a点不兴奋,d 点可兴奋,电流计只发生一次偏转。
二、兴奋的传导方向、特点的分析判断与设计
1.兴奋在完整反射弧中的传导方向判断与分析由于兴奋在神经元之间的传递是单向的,导致兴奋在完整反射弧中的传导也是单向的,只能由传入神经传人,由传出神经传出。
2.兴奋传导特点的设计验证
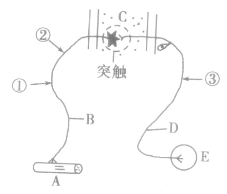
(1)验证冲动在神经纤维上的传导方法设计:电刺激图①处,观察A的变化,同时测量②处的电位有无变化。结果分析:若A有反应,且②处电位改变,说明冲动在神经纤维上的传导是双向的;若A有反应而②处无电位变化,则说明冲动在神经纤维上的传导是单向的。
(2)验证冲动在神经元之间的传递方法设计:先电刺激图①处,测量③处电位变化:再电刺激③处,测量①处的电位变化。结果分析:若两次实验的检测部位均发生电位变化,说明冲动在神经元间的传递是双向的;若只有一处电位改变,则说明冲动在神经元间的传递是单向的。
与“对下列四幅图的描述,正确的是[ ]A.图甲可依次表示果蝇的一...”考查相似的试题有:
- 果蝇中,正常翅(A)对短翅(a)为显性,此对等位基因位于常染色体上;红眼(B)对白眼(b)为显性,此对等位基因位于X染色体上。现有...
- 下图甲乙是某一生物个体(2n=4)的细胞分裂示意图,图丙是另一生物部分染色体分裂图像。请回答以下问题:(1)图甲所处的细胞分裂...
- 豌豆的红花对白花是显性,长花粉对圆花粉是显性。现有红花长花粉与白花圜花粉植株杂交,F1都是红花长花粉。若F1自交获得200株...
- 紫色种皮、厚壳花生和红色种皮、薄壳花生杂交,F1全是紫皮、厚壳花生,自交产生F2,F2中杂合紫皮、薄壳花生有3966株,问纯合...
- 以下二倍体生物的细胞中含有两个染色体组的是①有丝分裂中期细胞 ②有丝分裂后期细胞 ③减数第一次分裂中期细胞 ④减数第二次分裂...
- 下图中甲~丁为小鼠睾丸中细胞分裂不同时期的染色体数、染色单体数和DNA分子数的比例图,关于此图叙述中错误的是[ ]A.甲图可...
- 下图所表示的4个细胞中,只含有一个染色体组的是[ ]A.B.C.D.
- 如图所示,图1为细胞膜亚显微结构示意图,图2为突触结构示意图,则相关叙述正确的是[ ]A.图1中I侧为细胞膜内侧,Ⅱ侧为细胞膜...
- 以下有关“神经元之间通过突触传递信息图解”的说法中,正确的是[ ]A.静息状态下,突触前膜和突触后膜的膜两侧电位均表现为内...
- 人在拔牙时,往往需要在相应部位注射局部麻醉药,使其感觉不到疼痛,这是因为麻醉药[ ]A.阻断了传入神经的兴奋传导B.抑制神...