本试题 “下列说法中不正确的是[ ]A.加热杀死的S型肺炎双球菌和活的R型肺炎双球菌的混合物可使健康小鼠死亡B.高等植物光合作用过程中ATP、[H]和O2都在叶绿体基粒中产...” 主要考查您对光合作用的过程
遗传物质
DNA分子的结构
染色体变异
等考点的理解。关于这些考点您可以点击下面的选项卡查看详细档案。
- 光合作用的过程
- 遗传物质
- DNA分子的结构
- 染色体变异
光合作用过程:
1、光合作用的概念:
绿色植物通过叶绿体,利用光能,把二氧化碳和水转化成储存着能量的有机物,并且释放出氧气的过程。
2、光合作用图解: 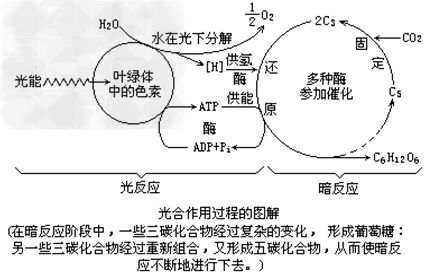
3、光合作用的总反应式及各元素去向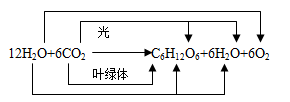
光反应与暗反应的比较:
| 项目 | 光反应(准备阶段) | 暗反应(完成阶段) |
| 场所 | 叶绿体的类囊体薄膜上 | 叶绿体的基质中 |
| 条件 | 光、色素、酶、水、ADP、 Pi |
多种酶、[H]、ATP、CO2、C5 |
| 物质变化 |  |
 |
| 能量的变化 | 光能转变成ATP中活跃的化学能 | ATP中活跃的化学能转变成(CH2O)中稳定的化学能 |
| 相互联系 | 光反应产物[H]、ATP为暗反应提供还原剂和能量;暗反应产生的ADP、Pi为光反应形成ATP提供了原料 | |
易错点拨:
1、光合作用总反应式两边的水不可轻易约去,因为反应物中的水在光反应阶段消耗,而产物中的水则在暗反应阶段产生。
2、催化光反应与暗反应的酶的分布场所不同,前者分布在类囊体薄膜上,后者分布在叶绿体基质中。
知识拓展:
1、氮能够提高光合作用的效率的原因是:氮是许多种酶的组成成分光合作用的场所:光合作用第一个阶段中的化学反应,必须有光才能进行。在类囊体的薄膜上进行;光合作用的第二个阶段中的化学反应,有没有光都可以进行。在叶绿体基质中进行。
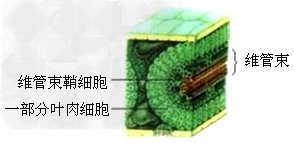
2、玉米是C4植物,其维管束鞘细胞中含有没有基粒的叶绿体,能够进行光合作用的暗反应。C4植物主要是那些生活在干旱热带地区的植物。
①四碳植物能利用强日光下产生的ATP推动PEP与CO2的结合,提高强光、高温下的光合速率,在干旱时可以部分地收缩气孔孔径,减少蒸腾失水,而光合速率降低的程度就相对较小,从而提高了水分在四碳植物中的利用率。
②二氧化碳固定效率比C3高很多,有利于植物在干旱环境生长。C3植物行光合作用所得的淀粉会贮存在叶肉细胞中;而C4植物的淀粉将会贮存于维管束鞘细胞内,维管束鞘细胞不含叶绿体。
3、光合细菌:利用光能和二氧化碳维持自养生活的有色细菌。光合细菌(简称PSB)是地球上出现最早、自然界中普遍存在、具有原始光能合成体系的原核生物,是在厌氧条件下进行不放氧光合作用的细菌的总称,是一类没有形成芽孢能力的革兰氏阴性菌,是一类以光作为能源、能在厌氧光照或好氧黑暗条件下利用自然界中的有机物、硫化物、氨等作为供氢体兼碳源进行光合作用的微生物。光合细菌广泛分布于自然界的土壤、水田、沼泽、湖泊、江海等处,主要分布于水生环境中光线能透射到的缺氧区。
1、遗传物质:
①概念:能单独传递遗传信息的物质。
②遗传物质的主要载体是染色体。
③作为遗传物质应具备的特点是:
a、分子结构具有相对稳定性;
b、能自我复制,保持上下代连续性;
c、能指导蛋白质合成;
d、能产生可遗传变异。
2、实验:包括肺炎双球菌转化实验、艾弗里证明DNA是遗传物质的实验(肺炎双球菌体外转化实验)、T2噬菌体侵染细菌的实验(用分别含有放射性同位素35S和放射性同位素32P培养基培养大肠杆菌。)、烟草花叶病毒的感染和重建实验。
实验证明DNA是主要的遗传物质,少部分生物的遗传物质是RNA。
(1)肺炎双球菌转化实验:
①肺炎双球菌
| S型细菌 | R型细菌 | |
| 菌落 | 光滑 | 粗糙 |
| 菌体 | 有多糖类荚膜 | 无多糖类荚膜 |
| 毒性 | 有毒性,使小鼠患败血症死亡 | 无毒性 |
a、研究者:1928年,英国科学家格里菲思。
b、实验材料:S型和R型肺炎双球菌、小鼠。
c、实验原理:S型肺炎双球菌使小鼠患败血病死亡;R型肺炎双球菌是无毒性的。
d、实验过程:
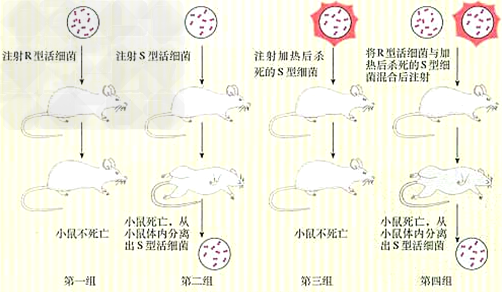
e、结论:加热杀死的S型细菌体内含有“转化因子”,促使R型细菌转化为S型细菌。
(2)艾弗里证明DNA是遗传物质的实验(肺炎双球菌体外转化实验):
a、研究者:1944年,美国科学家艾弗里等人。
b、实验材料:S型和R型肺炎双球菌、细菌培养基等。
c、实验设计思路:把DNA与其他物质分开,单独直接研究各自的遗传功能。
d、实验过程及分析
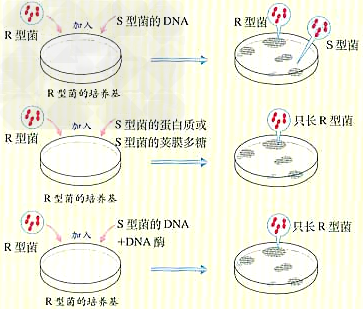
e、实验分析:①只有S型细菌的DNA能使R型细菌发生转化。 ②DNA被水解后不能使R型细菌发生转化。
d、实验结论:①S型细菌的DNA是“转化因子”,即DNA是遗传物质。 ②同时还直接证明蛋白质等其他物质不是遗传物质。
(3)T2噬菌体侵染细菌的实验:
a、研究着:1952年,赫尔希和蔡斯。
b、实验材料:T2噬菌体和大肠杆菌等。
c、实验方法:放射性同位素标记法。
d、实验思路: S是蛋白质的特有元素,DNA分子中含有P,蛋白质中几乎不含有,用放射性同位素32P和放射性同位素35S分别标记DNA和蛋白质,直接单独去观察它们的作用。
e、实验过程:标记细菌→标记噬菌体→用标记的噬菌体侵染普通细菌→搅拌离心。
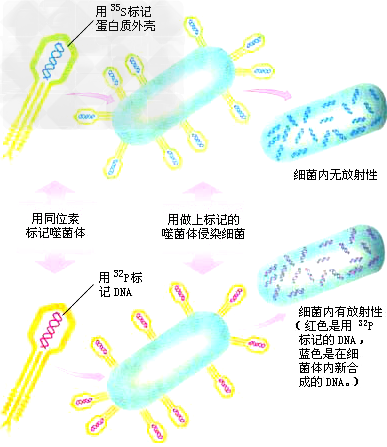
科学家首先用放射性同位素35S标记了一部分噬菌体的蛋白质,并用放射性同位素32P标记了另一部分噬菌体的DNA,然后,用被标记的T2噬菌体分别去侵染细菌(上图),当噬菌体在细菌体内大量增殖时,生物学家对被标记物质进行测试。
f、测试的结果表明,噬菌体的蛋白质并没有进入细菌内部,而是留在细菌的外部,噬菌体的DNA却进入了细菌体内,可见,噬菌体在细菌内的增殖是在噬菌体DNA的作用下完成的。即结论:在噬菌体中,亲代和子代间具有连续性的物质是DNA,即子代噬菌体的各种性状是通过亲代 DNA传给后代的,DNA才是真正的遗传物质。
体内转化实验与体外转化实验的比较:
| 体内转化实验 | 体外转化实验 | |
| 实验者 | 格里菲思 | 艾弗里及同事 |
| 培养细菌 | 用小鼠(体内) | 用培养基(体外) |
| 实验原则 | R型细菌与S型细菌的毒性对照 | S型细菌各成分作用的相互对照 |
| 实验结果 | 加热杀死的S型细菌能使R型细细菌转化为S型细菌 | S型细菌的DNA使R型菌转化为S型细菌 |
| 实验结论 | S型细菌体内有“转化因子” | S型细菌的DNA是遗传 物质 |
| 两实验联系: |
(1)所用材料相同,都是肺炎双球菌R型和S型 | |
肺炎双球菌体外转化实验和噬菌体侵染细菌实验的比较:
1.实验设计思路比较
| 艾弗里实验 | 噬菌体侵染细菌实验 | |
| 思路相同 | 设法将DNA与其他物质分开,单独、直接研究它们各自不同的遗传功能 | |
| 处理方式有区别 | 直接分离:分离S型细菌的DNA、多糖、蛋白质等,分别与R型细菌混合培养 | 同位素标记法:分别标记DNA和蛋白质的特殊元素(32P和35S) |
2.两个实验遵循相同的实验设计原则——对照原则
3.实验结论比较
(1)肺炎双球菌转化实验的结论:证明DNA是遗传物质,蛋白质不是遗传物质。
(2)噬菌体侵染细菌实验的结论:证明DNA是遗传物质,不能证明蛋白质不是遗传物质,因为蛋白质没有进入细菌体内。
知识点拨:
1、上清液和沉淀物中都有放射性的原因分析:
①用32P标记的噬菌体侵染大肠杆菌,上清液中有少量放射性的原因:
a.保温时间过短,部分噬菌体没有侵染到大肠杆菌细胞内,经离心后分布于上清液中,使上清液出现放射陡。
b.保温时间过长,噬菌体在大肠杆菌内增殖后释放子代,经离心后分布于上清液中,也会使上清液的放射性含量升高。
②用35S标记的噬菌体侵染大肠杆菌,沉淀物中也有少量放射性的原因:由于搅拌不充分,有少量35S的噬菌体蛋白质外壳吸附在细菌表面,随细菌离心到沉淀物中,使沉淀物中出现少量的放射性。
2、关于噬菌体侵染细菌实验中放射性元素的去向问题
①若用32P和35S标记病毒而宿主细胞未被标记,相当于间接地将核酸和蛋白质分开,只在子代病毒的核酸中有32p标记。
②若用32p和35S标记宿主细胞而病毒未被标记,则在子代病毒的核酸和蛋白质外壳中均有标记元素。
③若用C、H、O、N等标记病毒而宿主细胞未被标记,则只在子代病毒的核酸中有标记元素。
④若用C、H、O、N等标记宿主细胞而病毒未被标记,则在子代病毒的核酸和蛋白质外壳中均可找到标记元素。
3、标记噬菌体时应先标记细菌,用噬菌体侵染被标记的细菌,这样来标记噬菌体。因为噬菌体是没有细胞结构的病毒,只能在宿主细胞中繁殖后代,所以在培养基中它是不能繁殖后代的。
4、噬菌体侵染细菌实验只能证明DNA是遗传物质,不能证明蛋白质不是遗传物质,因蛋白质没有进入细菌体内。除此之外,还能证明DNA能进行自我复制,DNA控制蛋白质的合成。
5、两个实验都不能证明DNA是主要的遗传物质。
6、细胞生物(包括原核和真核生物)含有两种核酸(DNA和RNA),遗传物质是DNA;病毒没有细胞结构,只有一种核酸(DNA病毒、RNA病毒)。
7、DNA有四种碱基(AGCT),四种脱氧核苷酸。RNA有四种碱基(AGCU),四种核糖核苷酸。
知识拓展:
1、实验设计的基本思路是设法把 DNA和蛋白质分开,单独观察它们的作用。
2、加热杀死的S型细菌,其蛋白质变性失活; DNA在加热过程中,双螺旋解开,氢键被打断,但缓慢冷却时,其结构可恢复。
3、转化因子的实质是S型细菌的DNA片段整合到了R型细菌的DNA中,即实现了基因重组。
4、T2噬菌体
(1)结构:
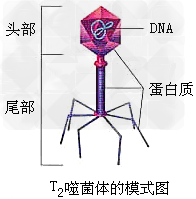
(2)T2噬菌体是一种专门寄生在大肠杆菌体内的病毒,它的头部和尾部的外壳都是由蛋白质构成的,头部内含有DNA。T2噬菌体侵染大肠杆菌后,就会在自身遗传物质的作用下,利用大肠杆菌体内的物质来合成自身的组成成分,进行大量增殖。当噬菌体增殖到一定数量后,大肠杆菌裂解,释放出大量的噬菌体。
5、侵染特点及过程
①进入细菌体内的是噬菌体的DNA,噬菌体的蛋白质外壳留在外面不起作用。
②噬菌体侵染细菌要经过吸附→注入核酸→合成 →组装→释放五个过程。
6、增殖特点:在自身遗传物质的作用下,利用大肠杆菌体内的物质合成自身成分,进行增殖。
DNA分子的结构:
1、DNA的元素组成:C、H、O、N、P
2、DNA分子的结构:DNA的双螺旋结构,两条反向平行脱氧核苷酸链,外侧磷酸和脱氧核糖交替连结,内侧碱基对(氢键)碱基互补配对原则。
3、模型图解: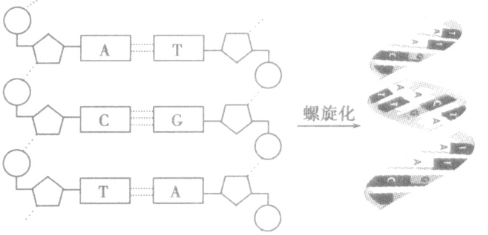
4、DNA分子的结构特性
(l)稳定性:DNA分子中脱氧核糖和磷酸交替连接的方式不变;两条链间碱基互补配对的方式不变。
(2)多样性:DNA分子中碱基时排列顺序多种多样。
(3)特异性:每种DNA有别于其他DNA的特定的碱基排列顺序。
知识点拨:
碱基互补配对的规律:

知识拓展:
1、两条链之间的脱氧核苷酸数目相等→两条链之间的碱基、脱氧核糖和磷酸数目对应相等。
2、碱基配对的关系是:A(或T)一定与T(或A)配对、G(或C)一定与C(或G)配对,这就是碱基互补配对原则。其中,A与T之间形成2个氢键,G与C之间形成3个氢键。
3、DNA分子彻底水解时得到的产物是脱氧核苷酸的基本组分,即脱氧核糖、磷酸、含氮碱基。
1、染色体变异分为染色体结构变异和数目变异。
(1)染色体结构变异
①概念:排列在染色体上的基因的数目或排列顺序发生改变,而导致性状的变异。
②类型:在自然条件或人为因素的影响下,染色体结构的变异主要有以下4种:缺失、重复、倒位、易位。 ③结果:染色体结构变异都会使排列在染色体上的基因的数目或排列顺序发生改变,从而导致性状的改变。
| 类型 | 定义 | 实例 | 示意图 |
| 缺失 | 一条正常染色体断裂后丢失某一片段引起的变异。 | 猫叫综合征 |  |
| 重复 | 染色体增加某一片段引起的变异。一条染色体的某一片段连接到同源的另一条染色体上,结果后者就有一段重复基因。 | 果蝇棒状眼 |  |
| 倒位 | 染色体中某一片段位置颠倒180°后重新结合到原部位引起的变异。基因并不丢失,因此一般生活正常。 | — |  |
| 易位 | 染色体的某一片段移接到另一条非同源染色体上引起的变异 | 人慢性粒细胞白血病 |  |
(1)染色体数目变异的种类
①细胞内的个别染色体增加或减少。
②细胞内染色体数日以染色体组的形式成倍地增加或减少,
(2)染色体组
①概念:细胞中的一组非同源染色体,它们在形态和功能上各不相同,但是携带着控制一种生物生长发育、遗传和变异的全部遗传信息,这样的一组染色体叫做一个染色体组。
②条件:
a、一个染色体组中不含有同源染色体;
b、一个染色体组中所含的染色体形态、大小和功能各不相同;
c、一个染色体组中含有控制生物性状的一整套基因。
(3)单倍体和多倍体比较
| 项目 | 单倍体 | 多倍体 | |
| 概念 | 体细胞中含有本物种配子染包体数目的个体 | 体细胞中含有三个或三个以上染色体组的个体 | |
| 成因 | 自然成因 | 由配子直接发育成个体,如雄蜂是由未受精的卵细胞发育而来 | 外界环境条件剧变 |
| 人工诱导 | 花药离体培养 | 用秋水仙素处理萌发的种子或幼苗 | |
| 发育起点 | 配子 | 受精卵或合子 | |
| 植株特点 | 植株弱小 | 茎秆粗壮,叶片、果实和种子比较大,营养物质含量丰富,发育延迟,结实率低 | |
| 可育性 | 高度不育 | 可育,但结实性差 | |
| 应用 | 单倍体育种 | 多倍体育种 | |
②染色体组:细胞中的一组非同源染色体,它们在形态和功能上各不相同,但是携带着控制一种生物生长发育、遗传和变异的全部信息,这样的一组染色体,叫做一个染色体组。
3、染色体变异在实践中的应用
(1)单倍体育种

例:
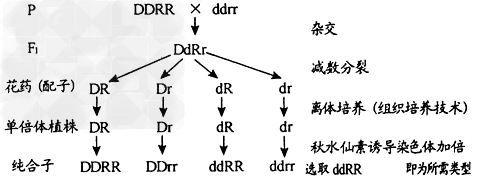
②优点:明显缩短育种年限,后代一般是纯合子。
(2)多倍体育种
①方法:用秋水仙素处理萌发的种子或幼苗。
②成因:秋水仙素抑制纺锤体的形成。
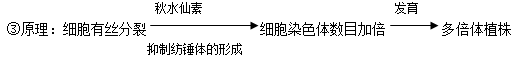
④实例:二倍体无子西瓜的培育。

表解基因重组、基因突变和染色体变异的不同:
| 项目 | 基因重组 | 基因突变 | 染色体变异 |
| 概念 | 因基因的重新组合而发生的变异 | 基因结构的改变,包括DNA碱基对的替换、增添和缺失 | 染色体结构或数目变化而引起的变异 |
| 类型 | ①非同源染色体上的非等位基因自由组合;②同源染色体的非姐妹染色单体之间的交叉互换 | ①自然状态下发生的——自然突变;②人为条件下发生的——人工诱变 | ①染色体结构变异;②染色体数目变异 |
| 鉴定方法 | 光学显微镜下均无法检出,可根据是否有新性状或新性状组合确定 | 光学显微镜下可检出 | |
| 适用范围 | 真核生物进行有性生殖的过程中发生 | 任何生物均可发生(包括原核生物、真核生物及非细胞结构的生物) | 真核生物遗传中发生 |
| 生殖类型 | 自然状态下只在有性生殖中发生 | 无性生殖和有性生殖均可发生 | 无性生殖和有性生殖均可发生 |
| 产生机理 | 由基因的自由组合和交叉互换引起 | 基因的分子结构发生改变的结果 | 染色体的结构或数目发生变化的结果 |
| 细胞分裂 | 在减数分裂中发生 | 无丝分裂、有丝分裂、减数分裂均可发生 | 有丝分裂和减数分裂中均可发生 |
| 产生结果 | 只改变基因型,未发生基因的改变,既无“质”的变化,也无“量”的变化 | 产生新的基因,发生基因“种类”的改变,即有“质”的变化,但无“量”的变化 | 可引起基因“数量”的变化,如增添或缺失几个基因 |
| 意义 | 生物变异的来源之一,对生物进化有十分重要的意义 | 生物变异的根本来源,提供生物进化的原材料 | 对生物进化有一定意义 |
| 育种应用 | 杂交育种 | 诱变育种 | 单倍体、多倍体育种 |
知识点拨:染色体组数的判定
1.染色体组数的判断方法
(1)根据细胞中染色体的形态判断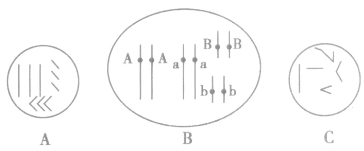
细胞内同一种形态的染色体有几条,则含有几个染色体组。如图A细胞内同种形态的染色体有3条,则该细胞中有3个染色体组;图C细胞内同一种形态的染色体有1条,则该细胞中有1个染色体组。
细胞内有几种形态的染色体,一个染色体组内就有几条染色体。如图A细胞内有3种形态的染色体,则该细胞的一个染色体组内就有3条染色体;如图C 细胞内有5种形态的染色体,则该细胞的一个染色体组内就有5条染色体。
(2)根据基因型判断
在细胞或生物体的基因型中,控制同一性状的基因出现几次,则含有几个染色体组,可简记为“同一英文字母无论大写还是小写,出现几次就含几个染色体组”。如图B细胞内控制同一性状的基因出现4次,则含有4个染色体组。
(3)根据染色体数目的形态数判断
染色体组的数目=染色体数/染色体的形态数
如图A细胞内共含有9条染色体,染色体的形态数是3种,9/3=3,则该细胞内含有3个染色体组;如图 B细胞内共含有8条染色体,染色体的形态数是2种, 8/2=4,则该细脆内含有4个染色体组;如图C细胞内共含有5条染色体,染色体的形态数是5种,5/5=1,则该细胞内含有1个染色体组。
2.一些细胞分裂图中的染色体组数判断(如图)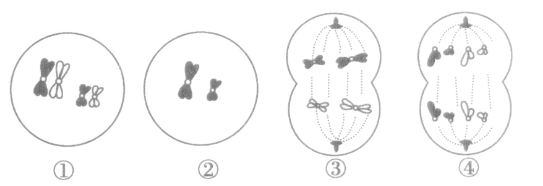
①减数第一次分裂的前期,染色体4条,生殖细胞中含有染色体2条,每个染色体组有2条染色体,该细胞中有2个染色体组。
②减数第二次分裂的前期,染色体2条,生殖细胞中含有染色体2条,每个染色体组有2条染色体,该细胞中有1个染色体组。
③减数第一次分裂的后期,染色体4条,生殖细胞中含有染色体2条,每个染色体组有、2条染色体,该细胞中有2个染色体组。
④有丝分裂后期,染色体8条,生殖细胞中含有染色体2条,每个染色体组有2条染色体,该细胞中有4 个染色体组。
知识拓展:
1、基因突变是染色体的某一位点上基因中碱基对的改变,是分子水平的变异,而染色体变异则是比较明显的染色体结构或数目的变异,属于细胞水平的变异。
2、判定生物是单倍体、二倍体、多倍体的关键是看它的发育起点。若发育起点是配子,不论其体细胞中含有几个染色体组都叫单倍体。若发育起点是受精卵,其体细胞中有几个染色体组就叫几倍体。
3、体细胞染色体组为奇数的单倍体与多倍体高度不育的原因:进行减数分裂形成配子时,同源染色体无法正常联会或联会紊乱,不能产生正常配子。
4、单倍体育种得到的一般是纯合子。二倍体生物的花粉经单倍体育种后,得到的一定是纯合子植株。四倍体等多倍体的花粉经离体培养、秋水仙素处理后,可能产生杂合子。如BBbb的花粉基因型有三种:BB、 Bb、bb,培养处理后基因型分别是BBBB、BBbb(杂合子)、bbbb。
5、用秋水仙素处理使植株染色体数目加倍,若操作对象是单倍体植株,叫单倍体育种;若操作对象为正常植株,叫多倍体育种。不能看到“染色体数目加倍” 就认为是多倍体育种。
6、不同生物的变异类型不同,不同生殖方式所带来的变异类型亦不相同,探究变异原因与变异类型时首先应注意的是生物的不同种类和生殖方式。
(1)病毒的可遗传变异的来源——基因突变。
(2)原核生物可遗传变异的来源——基因突变。
(3)真核生物可遗传变异的来源:
①进行无性生殖时——基因突变和染色体变异;
②进行有性生殖时——基因突变、基因重组和染色体变异。
与“下列说法中不正确的是[ ]A.加热杀死的S型肺炎双球菌和活的R...”考查相似的试题有:
- 利用同位素作为示踪元素,标记特定的化合物以追踪物质运行和变化过程的方法叫做同位素标记法。下列各项所选择使用的同位素及...
- 下列关于植物光合作用和细胞呼吸的叙述,正确的是[ ]A.在一定范围内,温度越高,光合速率和呼吸速率就越大B.在一定光照强度...
- 下列叙述正确的是[ ]A.DNA是蛋白质合成的直接模板B.每种氨基酸仅由一种密码子编码C.DNA复制就是基因表达的过程D.DNA是主...
- 噬菌体在细菌内合成自己的蛋白质需要[ ]A.噬菌体的DNA和氨基酸B.噬菌体的DNA和细菌的氨基酸C.细菌的DNA和氨基酸D.细菌的D...
- 为研究转化因子的物质属性,美科学家艾弗里等从S型细菌中提取DNA 、蛋白质、多糖等,然后将它们分别加入培养R型细菌的培养基...
- 艾弗里的肺炎双球菌转化实验和赫尔希、蔡斯的噬菌体侵染细菌实验,都能证明DNA是遗传物质,对这两个实验的研究方法:①设法把D...
- 某蛋白质由65个氨基酸组成,控制合成该蛋白质的基因的碱基数目至少有[ ]A.390个B.195个C.65个D.260个
- DNA的一条单链中=0.8,上述比例在其互补链和整个DNA分子中分别是[ ]A.0.8,1.0B.1.25,1.0C.0.8,0.8D.0.2,1.0
- 下图为生物遗传信息表达的示意图,请据图回答:(1)图中①过程在细胞的________内完成。(2)图中②表示________过程,进行的场所...
- DNA的一个单链中(A+G)/(T+C)=0.2,则(A+G)/(T+C)在其互补链和整个DNA分子中分别是[ ]A.0.4和0.6B.5和1.0C.0.4和0.4D.0.6和1.0