本试题 “以下是科学家在研究生物体的遗传和变异时做过的一些实验。根据实验分析回答:(1)实验1:用35S和32P分别标记噬菌体的蛋白质和DNA后,侵染未被标记的细菌,得到...” 主要考查您对生物的性状
遗传物质
染色体变异
科学研究方法
等考点的理解。关于这些考点您可以点击下面的选项卡查看详细档案。
- 生物的性状
- 遗传物质
- 染色体变异
- 科学研究方法
1、生物性状:生理方面的特征,形态方面的特征和行为方式的特征。
2、性状类型:
(1)相对性状:一种生物的同一种性状的不同表现类型。
(2)性状分离:杂种后代中,同时出现显性性状和隐性性状的现象。
如在DD×dd杂交实验中,杂合F1代自交后形成的F2代同时出现显性性状(DD及Dd)和隐性性状(dd)的现象。
(3)显性性状:在DD×dd杂交试验中,F1表现出来的性状;
如教材中F1代豌豆表现出高茎,即高茎为显性。决定显性性状的为显性遗传因子(基因),用大写字母表示。如高茎用D表示。
(4)隐性性状:在DD×dd杂交试验中,F1未显现出来的性状;
如教材中F1代豌豆未表现出矮茎,即矮茎为隐性。决定隐性性状的为隐性基因,用小写字母表示,如矮茎用d表示。等位基因:控制相对性状的基因。
(5)显性相对性:具有相同性状的亲本杂交,杂种子一代中不分显隐性,表现出两者的中间性状(不完全显性)或者是同事表现出两个亲本的性状(共显性)。
知识点拨:
1、生物的性状表现是基因型与环境相互作用的结果。
2、生物性状的鉴定:
①鉴定一只白羊是否纯合——测交
②在一对相对性状中区分显隐性——杂交
③不断提高小麦抗病品种的纯合度——自交
④检验杂种F1的基因型——测交
1、遗传物质:
①概念:能单独传递遗传信息的物质。
②遗传物质的主要载体是染色体。
③作为遗传物质应具备的特点是:
a、分子结构具有相对稳定性;
b、能自我复制,保持上下代连续性;
c、能指导蛋白质合成;
d、能产生可遗传变异。
2、实验:包括肺炎双球菌转化实验、艾弗里证明DNA是遗传物质的实验(肺炎双球菌体外转化实验)、T2噬菌体侵染细菌的实验(用分别含有放射性同位素35S和放射性同位素32P培养基培养大肠杆菌。)、烟草花叶病毒的感染和重建实验。
实验证明DNA是主要的遗传物质,少部分生物的遗传物质是RNA。
(1)肺炎双球菌转化实验:
①肺炎双球菌
| S型细菌 | R型细菌 | |
| 菌落 | 光滑 | 粗糙 |
| 菌体 | 有多糖类荚膜 | 无多糖类荚膜 |
| 毒性 | 有毒性,使小鼠患败血症死亡 | 无毒性 |
a、研究者:1928年,英国科学家格里菲思。
b、实验材料:S型和R型肺炎双球菌、小鼠。
c、实验原理:S型肺炎双球菌使小鼠患败血病死亡;R型肺炎双球菌是无毒性的。
d、实验过程:
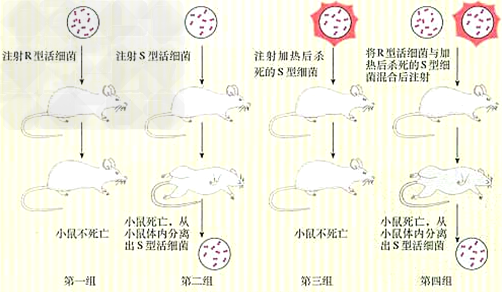
e、结论:加热杀死的S型细菌体内含有“转化因子”,促使R型细菌转化为S型细菌。
(2)艾弗里证明DNA是遗传物质的实验(肺炎双球菌体外转化实验):
a、研究者:1944年,美国科学家艾弗里等人。
b、实验材料:S型和R型肺炎双球菌、细菌培养基等。
c、实验设计思路:把DNA与其他物质分开,单独直接研究各自的遗传功能。
d、实验过程及分析
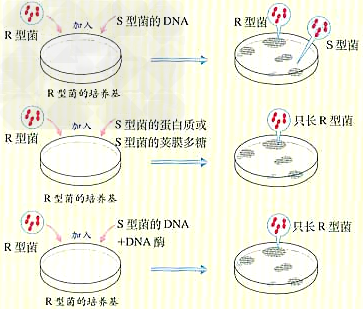
e、实验分析:①只有S型细菌的DNA能使R型细菌发生转化。 ②DNA被水解后不能使R型细菌发生转化。
d、实验结论:①S型细菌的DNA是“转化因子”,即DNA是遗传物质。 ②同时还直接证明蛋白质等其他物质不是遗传物质。
(3)T2噬菌体侵染细菌的实验:
a、研究着:1952年,赫尔希和蔡斯。
b、实验材料:T2噬菌体和大肠杆菌等。
c、实验方法:放射性同位素标记法。
d、实验思路: S是蛋白质的特有元素,DNA分子中含有P,蛋白质中几乎不含有,用放射性同位素32P和放射性同位素35S分别标记DNA和蛋白质,直接单独去观察它们的作用。
e、实验过程:标记细菌→标记噬菌体→用标记的噬菌体侵染普通细菌→搅拌离心。
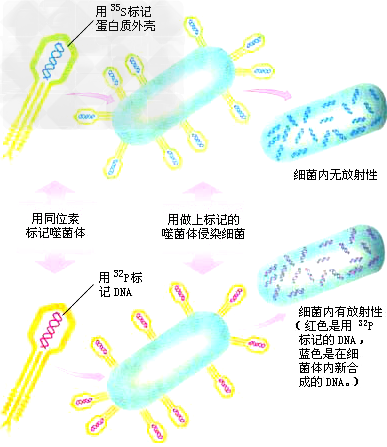
科学家首先用放射性同位素35S标记了一部分噬菌体的蛋白质,并用放射性同位素32P标记了另一部分噬菌体的DNA,然后,用被标记的T2噬菌体分别去侵染细菌(上图),当噬菌体在细菌体内大量增殖时,生物学家对被标记物质进行测试。
f、测试的结果表明,噬菌体的蛋白质并没有进入细菌内部,而是留在细菌的外部,噬菌体的DNA却进入了细菌体内,可见,噬菌体在细菌内的增殖是在噬菌体DNA的作用下完成的。即结论:在噬菌体中,亲代和子代间具有连续性的物质是DNA,即子代噬菌体的各种性状是通过亲代 DNA传给后代的,DNA才是真正的遗传物质。
体内转化实验与体外转化实验的比较:
| 体内转化实验 | 体外转化实验 | |
| 实验者 | 格里菲思 | 艾弗里及同事 |
| 培养细菌 | 用小鼠(体内) | 用培养基(体外) |
| 实验原则 | R型细菌与S型细菌的毒性对照 | S型细菌各成分作用的相互对照 |
| 实验结果 | 加热杀死的S型细菌能使R型细细菌转化为S型细菌 | S型细菌的DNA使R型菌转化为S型细菌 |
| 实验结论 | S型细菌体内有“转化因子” | S型细菌的DNA是遗传 物质 |
| 两实验联系: |
(1)所用材料相同,都是肺炎双球菌R型和S型 | |
肺炎双球菌体外转化实验和噬菌体侵染细菌实验的比较:
1.实验设计思路比较
| 艾弗里实验 | 噬菌体侵染细菌实验 | |
| 思路相同 | 设法将DNA与其他物质分开,单独、直接研究它们各自不同的遗传功能 | |
| 处理方式有区别 | 直接分离:分离S型细菌的DNA、多糖、蛋白质等,分别与R型细菌混合培养 | 同位素标记法:分别标记DNA和蛋白质的特殊元素(32P和35S) |
2.两个实验遵循相同的实验设计原则——对照原则
3.实验结论比较
(1)肺炎双球菌转化实验的结论:证明DNA是遗传物质,蛋白质不是遗传物质。
(2)噬菌体侵染细菌实验的结论:证明DNA是遗传物质,不能证明蛋白质不是遗传物质,因为蛋白质没有进入细菌体内。
知识点拨:
1、上清液和沉淀物中都有放射性的原因分析:
①用32P标记的噬菌体侵染大肠杆菌,上清液中有少量放射性的原因:
a.保温时间过短,部分噬菌体没有侵染到大肠杆菌细胞内,经离心后分布于上清液中,使上清液出现放射陡。
b.保温时间过长,噬菌体在大肠杆菌内增殖后释放子代,经离心后分布于上清液中,也会使上清液的放射性含量升高。
②用35S标记的噬菌体侵染大肠杆菌,沉淀物中也有少量放射性的原因:由于搅拌不充分,有少量35S的噬菌体蛋白质外壳吸附在细菌表面,随细菌离心到沉淀物中,使沉淀物中出现少量的放射性。
2、关于噬菌体侵染细菌实验中放射性元素的去向问题
①若用32P和35S标记病毒而宿主细胞未被标记,相当于间接地将核酸和蛋白质分开,只在子代病毒的核酸中有32p标记。
②若用32p和35S标记宿主细胞而病毒未被标记,则在子代病毒的核酸和蛋白质外壳中均有标记元素。
③若用C、H、O、N等标记病毒而宿主细胞未被标记,则只在子代病毒的核酸中有标记元素。
④若用C、H、O、N等标记宿主细胞而病毒未被标记,则在子代病毒的核酸和蛋白质外壳中均可找到标记元素。
3、标记噬菌体时应先标记细菌,用噬菌体侵染被标记的细菌,这样来标记噬菌体。因为噬菌体是没有细胞结构的病毒,只能在宿主细胞中繁殖后代,所以在培养基中它是不能繁殖后代的。
4、噬菌体侵染细菌实验只能证明DNA是遗传物质,不能证明蛋白质不是遗传物质,因蛋白质没有进入细菌体内。除此之外,还能证明DNA能进行自我复制,DNA控制蛋白质的合成。
5、两个实验都不能证明DNA是主要的遗传物质。
6、细胞生物(包括原核和真核生物)含有两种核酸(DNA和RNA),遗传物质是DNA;病毒没有细胞结构,只有一种核酸(DNA病毒、RNA病毒)。
7、DNA有四种碱基(AGCT),四种脱氧核苷酸。RNA有四种碱基(AGCU),四种核糖核苷酸。
知识拓展:
1、实验设计的基本思路是设法把 DNA和蛋白质分开,单独观察它们的作用。
2、加热杀死的S型细菌,其蛋白质变性失活; DNA在加热过程中,双螺旋解开,氢键被打断,但缓慢冷却时,其结构可恢复。
3、转化因子的实质是S型细菌的DNA片段整合到了R型细菌的DNA中,即实现了基因重组。
4、T2噬菌体
(1)结构:
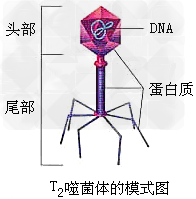
(2)T2噬菌体是一种专门寄生在大肠杆菌体内的病毒,它的头部和尾部的外壳都是由蛋白质构成的,头部内含有DNA。T2噬菌体侵染大肠杆菌后,就会在自身遗传物质的作用下,利用大肠杆菌体内的物质来合成自身的组成成分,进行大量增殖。当噬菌体增殖到一定数量后,大肠杆菌裂解,释放出大量的噬菌体。
5、侵染特点及过程
①进入细菌体内的是噬菌体的DNA,噬菌体的蛋白质外壳留在外面不起作用。
②噬菌体侵染细菌要经过吸附→注入核酸→合成 →组装→释放五个过程。
6、增殖特点:在自身遗传物质的作用下,利用大肠杆菌体内的物质合成自身成分,进行增殖。
1、染色体变异分为染色体结构变异和数目变异。
(1)染色体结构变异
①概念:排列在染色体上的基因的数目或排列顺序发生改变,而导致性状的变异。
②类型:在自然条件或人为因素的影响下,染色体结构的变异主要有以下4种:缺失、重复、倒位、易位。 ③结果:染色体结构变异都会使排列在染色体上的基因的数目或排列顺序发生改变,从而导致性状的改变。
| 类型 | 定义 | 实例 | 示意图 |
| 缺失 | 一条正常染色体断裂后丢失某一片段引起的变异。 | 猫叫综合征 |  |
| 重复 | 染色体增加某一片段引起的变异。一条染色体的某一片段连接到同源的另一条染色体上,结果后者就有一段重复基因。 | 果蝇棒状眼 |  |
| 倒位 | 染色体中某一片段位置颠倒180°后重新结合到原部位引起的变异。基因并不丢失,因此一般生活正常。 | — |  |
| 易位 | 染色体的某一片段移接到另一条非同源染色体上引起的变异 | 人慢性粒细胞白血病 |  |
(1)染色体数目变异的种类
①细胞内的个别染色体增加或减少。
②细胞内染色体数日以染色体组的形式成倍地增加或减少,
(2)染色体组
①概念:细胞中的一组非同源染色体,它们在形态和功能上各不相同,但是携带着控制一种生物生长发育、遗传和变异的全部遗传信息,这样的一组染色体叫做一个染色体组。
②条件:
a、一个染色体组中不含有同源染色体;
b、一个染色体组中所含的染色体形态、大小和功能各不相同;
c、一个染色体组中含有控制生物性状的一整套基因。
(3)单倍体和多倍体比较
| 项目 | 单倍体 | 多倍体 | |
| 概念 | 体细胞中含有本物种配子染包体数目的个体 | 体细胞中含有三个或三个以上染色体组的个体 | |
| 成因 | 自然成因 | 由配子直接发育成个体,如雄蜂是由未受精的卵细胞发育而来 | 外界环境条件剧变 |
| 人工诱导 | 花药离体培养 | 用秋水仙素处理萌发的种子或幼苗 | |
| 发育起点 | 配子 | 受精卵或合子 | |
| 植株特点 | 植株弱小 | 茎秆粗壮,叶片、果实和种子比较大,营养物质含量丰富,发育延迟,结实率低 | |
| 可育性 | 高度不育 | 可育,但结实性差 | |
| 应用 | 单倍体育种 | 多倍体育种 | |
②染色体组:细胞中的一组非同源染色体,它们在形态和功能上各不相同,但是携带着控制一种生物生长发育、遗传和变异的全部信息,这样的一组染色体,叫做一个染色体组。
3、染色体变异在实践中的应用
(1)单倍体育种

例:
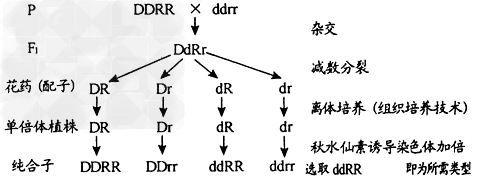
②优点:明显缩短育种年限,后代一般是纯合子。
(2)多倍体育种
①方法:用秋水仙素处理萌发的种子或幼苗。
②成因:秋水仙素抑制纺锤体的形成。
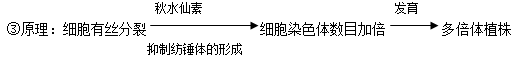
④实例:二倍体无子西瓜的培育。

表解基因重组、基因突变和染色体变异的不同:
| 项目 | 基因重组 | 基因突变 | 染色体变异 |
| 概念 | 因基因的重新组合而发生的变异 | 基因结构的改变,包括DNA碱基对的替换、增添和缺失 | 染色体结构或数目变化而引起的变异 |
| 类型 | ①非同源染色体上的非等位基因自由组合;②同源染色体的非姐妹染色单体之间的交叉互换 | ①自然状态下发生的——自然突变;②人为条件下发生的——人工诱变 | ①染色体结构变异;②染色体数目变异 |
| 鉴定方法 | 光学显微镜下均无法检出,可根据是否有新性状或新性状组合确定 | 光学显微镜下可检出 | |
| 适用范围 | 真核生物进行有性生殖的过程中发生 | 任何生物均可发生(包括原核生物、真核生物及非细胞结构的生物) | 真核生物遗传中发生 |
| 生殖类型 | 自然状态下只在有性生殖中发生 | 无性生殖和有性生殖均可发生 | 无性生殖和有性生殖均可发生 |
| 产生机理 | 由基因的自由组合和交叉互换引起 | 基因的分子结构发生改变的结果 | 染色体的结构或数目发生变化的结果 |
| 细胞分裂 | 在减数分裂中发生 | 无丝分裂、有丝分裂、减数分裂均可发生 | 有丝分裂和减数分裂中均可发生 |
| 产生结果 | 只改变基因型,未发生基因的改变,既无“质”的变化,也无“量”的变化 | 产生新的基因,发生基因“种类”的改变,即有“质”的变化,但无“量”的变化 | 可引起基因“数量”的变化,如增添或缺失几个基因 |
| 意义 | 生物变异的来源之一,对生物进化有十分重要的意义 | 生物变异的根本来源,提供生物进化的原材料 | 对生物进化有一定意义 |
| 育种应用 | 杂交育种 | 诱变育种 | 单倍体、多倍体育种 |
知识点拨:染色体组数的判定
1.染色体组数的判断方法
(1)根据细胞中染色体的形态判断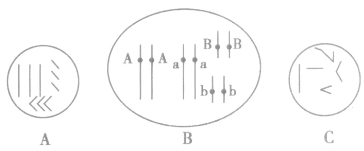
细胞内同一种形态的染色体有几条,则含有几个染色体组。如图A细胞内同种形态的染色体有3条,则该细胞中有3个染色体组;图C细胞内同一种形态的染色体有1条,则该细胞中有1个染色体组。
细胞内有几种形态的染色体,一个染色体组内就有几条染色体。如图A细胞内有3种形态的染色体,则该细胞的一个染色体组内就有3条染色体;如图C 细胞内有5种形态的染色体,则该细胞的一个染色体组内就有5条染色体。
(2)根据基因型判断
在细胞或生物体的基因型中,控制同一性状的基因出现几次,则含有几个染色体组,可简记为“同一英文字母无论大写还是小写,出现几次就含几个染色体组”。如图B细胞内控制同一性状的基因出现4次,则含有4个染色体组。
(3)根据染色体数目的形态数判断
染色体组的数目=染色体数/染色体的形态数
如图A细胞内共含有9条染色体,染色体的形态数是3种,9/3=3,则该细胞内含有3个染色体组;如图 B细胞内共含有8条染色体,染色体的形态数是2种, 8/2=4,则该细脆内含有4个染色体组;如图C细胞内共含有5条染色体,染色体的形态数是5种,5/5=1,则该细胞内含有1个染色体组。
2.一些细胞分裂图中的染色体组数判断(如图)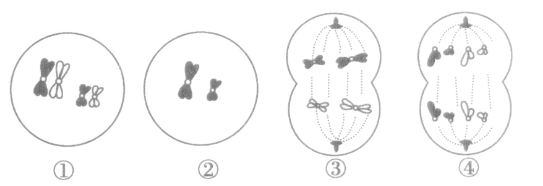
①减数第一次分裂的前期,染色体4条,生殖细胞中含有染色体2条,每个染色体组有2条染色体,该细胞中有2个染色体组。
②减数第二次分裂的前期,染色体2条,生殖细胞中含有染色体2条,每个染色体组有2条染色体,该细胞中有1个染色体组。
③减数第一次分裂的后期,染色体4条,生殖细胞中含有染色体2条,每个染色体组有、2条染色体,该细胞中有2个染色体组。
④有丝分裂后期,染色体8条,生殖细胞中含有染色体2条,每个染色体组有2条染色体,该细胞中有4 个染色体组。
知识拓展:
1、基因突变是染色体的某一位点上基因中碱基对的改变,是分子水平的变异,而染色体变异则是比较明显的染色体结构或数目的变异,属于细胞水平的变异。
2、判定生物是单倍体、二倍体、多倍体的关键是看它的发育起点。若发育起点是配子,不论其体细胞中含有几个染色体组都叫单倍体。若发育起点是受精卵,其体细胞中有几个染色体组就叫几倍体。
3、体细胞染色体组为奇数的单倍体与多倍体高度不育的原因:进行减数分裂形成配子时,同源染色体无法正常联会或联会紊乱,不能产生正常配子。
4、单倍体育种得到的一般是纯合子。二倍体生物的花粉经单倍体育种后,得到的一定是纯合子植株。四倍体等多倍体的花粉经离体培养、秋水仙素处理后,可能产生杂合子。如BBbb的花粉基因型有三种:BB、 Bb、bb,培养处理后基因型分别是BBBB、BBbb(杂合子)、bbbb。
5、用秋水仙素处理使植株染色体数目加倍,若操作对象是单倍体植株,叫单倍体育种;若操作对象为正常植株,叫多倍体育种。不能看到“染色体数目加倍” 就认为是多倍体育种。
6、不同生物的变异类型不同,不同生殖方式所带来的变异类型亦不相同,探究变异原因与变异类型时首先应注意的是生物的不同种类和生殖方式。
(1)病毒的可遗传变异的来源——基因突变。
(2)原核生物可遗传变异的来源——基因突变。
(3)真核生物可遗传变异的来源:
①进行无性生殖时——基因突变和染色体变异;
②进行有性生殖时——基因突变、基因重组和染色体变异。
1、假说——演绎法
①提出假设
②演绎就是推理
③实验验证假设和推理
④得出结论
2、同位素示踪法:同位素示踪法是利用放射性核素或稀有稳定核素作为示踪剂对研究对象进行标记的微量分析方法
3、科学的研究方法包括:归纳法、类比推理法、实验法和演绎法。
①归纳法:是从个别性知识,引出一般性知识的推理,是由已知真的前提,引出可能真的结论。它把特性或关系归结到基于对特殊的代表(token)的有限观察的类型;或公式表达基于对反复再现的现象的模式(pattern)的有限观察的规律。
②类比推理法:类比推理这是科学研究中常用的方法之一。类比推理是根据两个或两类对象有部分属性相同,从而推出它们的其他属性也相同的推理。简称类推、类比。它是以关于两个事物某些属性相同的判断为前提,推出两个事物的其他属性相同的结论的推理。
③实验法:通过试验的论证得出所需数据,进行分析后得出结论。分为:化学物质的检测方法;实验结果的显示方法;实验条件的控制方法;实验中控制温度的方法
④演绎法:从普遍性结论或一般性事理推导出个别性结论的论证方法。演绎法得出的结论正确与否,有待于实践检验。它只能从逻辑上保证其结论的有效性,而不能从内容上确保其结论的真理性。也可以从逻辑思维,逆向思维和想象思维延伸到其结论该以反证明。
4、实验必须遵守的原则:
①设置对照原则:空白对照;条件对照;相互对照;自身对照。
②单一变量原则;
③平行重复原则
5、实验的特性:对照,统一性质。提出问题;设计方案;讨论结果;分析问题。分为科学实验;验证性实验;对照实验等。
知识拓展:
1、生物学的历史研究进展和相关实验的叙述。
(1)孟德尔的假说——演绎法叙述
①提出假设(如孟德尔根据亲本杂交实验,得到F1,Aa这对基因是独立的,在产生配子时相互分离。这里假设的是一对等位基因的情况);
②演绎就是推理(如果这个假说是正确的,这样F1会产生两种数量相等的配子,这样测交后代应该会产生两种数量相等的类型);
③最后实验验证假设和推理(测交实验验证,结果确实产生了两种数量相等的类型);
④最后得出结论(就是分离定律)
(2)遗传物质验证的三个实验:肺炎双球菌的转化实验;噬菌体侵染细菌的实验;烟草花叶病毒的重组实验
(3)酶发现过程中的实验:
①1777年,苏格兰医生史蒂文斯从胃里分离一种液体(胃液),并证明了食物的分解过程可以在体外进行。
②1834年,德国博物学家施旺把氯化汞加到胃液里,沉淀出一种白色粉末。除去粉末中的汞化合物,把剩下的粉末溶解,得到了一种浓度非常高的消化液,他把这粉末叫作“胃蛋白酶”(希腊语中的消化之意)。同时,两位法国化学家帕扬和佩索菲发现,麦芽提取物中有一种物质,能使淀粉变成糖,变化的速度超过了酸的作用,他们称这种物质为“淀粉酶制剂”(希腊语的“分离”)。科学家们把酵母细胞一类的活动体酵素和像胃蛋白酶一类的非活体酵素作了明确的区分。
③1878年,德国生理学家库恩提出把后者叫作“酶”。
④1897年,德国化学家毕希纳用砂粒研磨酵细胞,把所有的细胞全部研碎,并成功地提取出一种液体。他发现,这种液体依然能够像酵母细胞一样完成发酵任务。这个实验证明了活体酵素与非活体酵素的功能是一样的。因此,“酶”这个词现在适用于所有的酵素,而且是使生化反应的催化剂。由于这项发现,毕希纳获得了1907年诺贝尔化学奖
(4)生长素的发现实验:植物的向光生长和胚芽鞘实验
2、同位素示踪方法的应用,使人们可以从分子水平动态地观察生物体内或细胞内生理、生化过程,认识生命活动的物质基础。例如,用C、O等同位素研究光合作用,可以详细地阐明叶绿素如何利用二氧化碳和水,什么是从这些简单分子形成糖类等大分子的中间物,以及影响每步生物合成反应的条件等。
3、放射性同位素示踪技术,是分子生物学研究中的重要手段之一,对蛋白质生物合成的研究,从DNA复制、RNA转录到蛋白质翻译均起了很大的作用。最近邻序列分析法应用同位素示踪技术结合酶切理论和统计学理论,研究证实了DNA分子中碱基排列规律,在体外作合成DNA的实验:分四批进行,每批用一种不同的32P标记脱氧核苷三磷酸,32P标记在戊糖5'C的位置上,在完全条件下合成后,用特定的酶打开5'C-P键,使原碱基上通过戊糖5'C相连的32P移到最邻近的另一单核苷酸的3'C上。用最近邻序列分析法首次提出了DNA复制与RNA转录的分子生物学基础,从而建立了分子杂交技术,例如以噬体T2-DNA为模板制成[32P]RNA,取一定量T2-DNA和其它一些DNA加入此[32P]RNA中,经加热使DNA双链打开,并温育,用密度梯度离心或微孔膜分离出DNA-[32P]RNA复合体测其放射性,实验结果只有菌体T2的DNA能与该[32P]RNA形成放射性复合体。从而证明了RNA与DNA模板的碱基呈特殊配对的互补关系,用分子杂交技术还证实了从RNA到DNA的逆转录现象。
4、放射性同位素示踪技术对分子生物学的贡献还表现在:
a、对蛋白质合成过程中三个连续阶段,即肽链的起始、延伸和终止的研究;
b、核酸的分离和纯化;
c、核酸末端核苷酸分析,序列测定;
d、核酸结构与功能的关系;
e、RNA中的遗传信息如何通过核苷酸的排列顺序向蛋质中氨基酸传递的研究等等。
为了更好地应用放射性同位素示踪技术,除了有赖于示踪剂的高质量和核探测器的高灵敏度外,关键还在于有科学根据的设想和创造性的实验设计以及各种新技术的综合应用。
与“以下是科学家在研究生物体的遗传和变异时做过的一些实验。根...”考查相似的试题有:
- 现有两纯种小麦,一纯种小麦性状是高秆(D),抗锈病(T);另一纯种小麦的性状是矮秆(d),易染锈病(t)(两对基因独立遗传)。育种...
- 已知豌豆种子子叶的黄色与绿色是一对等位基因Y、 控制的,用豌豆进行下列遗传实验,具体情况如下:请分析回答:(1)从实验____...
- 紫茉莉的遗传信息储存于它的[ ]A.DNA或RNA分子,在细胞核与细胞质中B.DNA分子,在细胞核中C.DNA和RNA分子,在细胞核与细胞...
- 用32P标记S型肺炎双球菌的DNA,35S标记其蛋白质,将其加热杀死后与未作标记的R型活细菌混合并注入小鼠体内。一段时间后,从死...
- 在肺炎双球菌转化实验中,将R型活细菌与加热杀死的S型细菌混合后,注射到小鼠体内,下列能在死亡小鼠体内出现的细菌类型有①少...
- 20世纪50年代,科学家受达尔文进化思想的启发,广泛开展了人工动、植物育种研究,通过人工创造变异选育优良的新品种。这一过...
- 用甲物种农作物的①和②两个品种以及乙物种作原材料,分别培育出四个品种的过程如下图表示。请根据图回答问题:(1)①和②作为亲本...
- 给动物注射促甲状腺激素后最终导致下丘脑的促甲状腺激素释放激素分泌减少。对此问题的解释有两种观点:①促甲状腺激素直接对下...
- 实验一:选取同品种、同日龄的健康大鼠若干只,实施切除手术,一段时间后随机等分成四组,分别注射激素及生理盐水30天,结果...
- 为证明“唾液淀粉酶对淀粉有消化作用”,某同学制订了下列实验方案。(1)实验目的(略)(2)实验材料和用具(略)(3)实验方法和步骤①...